
Overview of the Equine Respiratory System
Read
The Upper Airway
The horse is an obligate nasal breather and all the air needed for gas exchange, at rest and during exercise, must pass through the upper airway. The upper airway provides a high resistance to airflow and may be a limiting factor in the horse’s exercise capacity [1,2]. For this reason alone, those interested in the athletic horse should have a thorough understanding of upper airway function. Furthermore, the upper airway is a common site of lesions limiting exercise capacity [3,4]. In this chapter we will review the upper airway as a conduit for airflow, discuss how the upper airway maintains patency during exercise, and describe quantitative measures of upper airway evaluation.
Upper Airway Flow Mechanics
A normal horse at rest breathing 20 times a minute with a tidal volume of 5 liters has a minute ventilation of 100 liters. As this horse begins to exercise, respiratory rate and tidal volume increase to achieve a minute ventilation of approximately 1,500 liters/minute [5,6]. The upper airway must accommodate this large increase in airflow by undergoing changes in caliber, rigidity, and shape. In spite of these adaptations to exercise, evidence suggests that in exercising horses upper airway resistance becomes a large portion of total respiratory resistance and that this increase in resistance may limit performance [2,7]. In strenuously exercising horses, the PaO2 decreases and the PaCO2 increases [8]. This perturbation in gas exchange is in part caused by insufficient alveolar ventilation, although diffusion limitation also plays a major role (see below). substituting a helium-oxygen mixture for air reduces airway resistance and blood oxygenation is improved [1]. Taken together, this is strong evidence to suggest that upper airway resistance limits arterial blood oxygenation in exercising horses, thereby limiting oxygen delivery to tissues and performance. Furthermore, if the function of the upper airway as a conduit for airflow is taxed to the maximum during exercise, it is easy to understand that small upper airway lesions may significantly impact upper airway function during exercise and adversely affect performance.
Distribution of Upper Airway Resistance
In the horse, as in most mammals, upper airway resistance is a significant portion of total resistance to flow. In the resting horse, a full two-thirds of the total resistance to airflow resides in the upper airway. This proportion increases during exercise [2]. Because of this, during exercise most animal species switch to mouth breathing. This provides a low-resistance pathway for the greater airflow required during exercise, thereby minimizing the work of breathing. Surprisingly, the horse cannot breathe through its mouth efficiently and the more than 20-fold increase in airflow that occurs during exercise must be totally accommodated by the upper airway. Obligate nasal breathing is the result of a tight seal between the soft palate and the laryngeal cartilages. When the nasal passages are occluded, this seal may be broken and mouth breathing results. However, in the horses, the resistance to airflow through the mouth is apparently very high.
As already mentioned, in the resting horse, upper airway resistance is a full two-thirds of the total resistance to airflow. Intrapulmonary airways from the trachea down to the small bronchioles contribute only one-third of the total resistance to flow in the standing horse. In the resting horse, nasal resistance increases slightly on inhalation and laryngeal resistance increases slightly on exhalation. During exercise, however, upper airway resistance changes greatly. In the exercising horse, during inhalation, intrathoracic resistance is less than 25% of total resistance to flow and the remaining resistance is almost evenly divided between nasal and laryngeal resistance. On exhalation, the intrathoracic resistance is about half the total resistance to flow, the other half being evenly divided between nasal and laryngeal resistance [2]. These changes in resistance during inhalation and exhalation can be explained by the pressure changes in the upper airway. In the resting horse, pressure changes are relatively modest and therefore changes in resistance are also small. On exhalation, there is positive pressure in the upper airway moving air out to the atmosphere. This positive pressure tends to dilate the upper airway, thereby decreasing resistance. Conversely, on inhalation, there is a negative pressure in the upper airway, relative to atmosphere. This is needed to move air in from the atmosphere via the upper airways into the lungs. This negative pressure tends to decrease airway caliber, thereby increasing resistance. During exercise, these pressure swings are greatly exaggerated because of the much higher airflows [5,9].
At first glance, it may be surprising that upper airway pressure swings are sufficient to cause such large changes in resistance. This phenomenon is explained by the observation that small changes in airway caliber cause large changes in resistance.
The following formula relates factors that determine the resistance of a cylindrical tube of length: L and radius of: r; with a fluid flow of a viscosity: u.
The formula is: R = 8 x u x L/r4
This equation demonstrates that radius is a very important determinant of resistance and that reducing the radius by half increases resistance 16-fold. Thus, a relatively small decrease in upper airway caliber can significantly increase resistance to flow and limit performance. Once the cross-sectional area of an airway region is decreased, air velocity increases and intra luminal pressure decreases (Bernoulli effect). This reduces airway cross-sectional area even further, setting up a vicious cycle resulting in airway collapse. This phenomenon is called dynamic collapse.
Upper airway caliber varies with region. Airway diameters are smallest at the external nares and larynx and largest through the nasal turbinate area and pharynx [10]. This observation has practical implications, because it suggests that lesions in high-resistance regions (external nares and larynx) are more likely to cause clinically significant upper airway obstruction than similar-sized lesions in regions such as the pharynx. For example, a small granulomatous lesion located on the axial surface of the arytenoid cartilage is of concern, because it reduces the airway caliber in a region of the upper airway that is already narrow. In contrast, pharyngeal lymphoid hyperplasia is not likely to significantly increase total resistance to air flow because this space-occupying lesion is located in the pharynx, which has a large cross-sectional area and therefore contributes less to total resistance to air flow.
Mechanisms Whereby the Upper Airway Withstands Changes in Caliber Induced by Changes in Pressure
Of course, the upper airway is designed to withstand the effects of pressure changes on its caliber. For much of its length, the upper airway is rigidly supported by bone and cartilage, while in other regions there is muscular support. Also, extension of the head and neck stiffens the airway and airway caliber can be increased by vasoconstriction of blood vessels in the mucosa.
Rigid Airway Support and the Laryngeal Mucosa
The most rigid support of the upper airway is by bone and cartilage. However, marked changes in airway caliber are possible even in these bone- and cartilage-supported regions. The upper airway is lined by mucosa, which contains capacitance vessels. These vessels are potentially very large and have the ability to fill with blood and thicken the upper airway mucosa to the point of occlusion. Normally, the size of capacitance vessels is kept small by tonic sympathetic innervation [11]. Injury to the sympathetic trunk innervating this region, for example in Horner’s syndrome, can cause complete nasal occlusion on the affected side [12]. Inflammatory disease of the nose can also cause mucosal enlargement by filling of capacitance vessels with blood [11]. The effects of common inflammatory diseases of horses, such as viral infections, on nasal patency have been insufficiently studied.
Muscular Support of the Upper Airway
In portions of the upper airway, such as the external nares, pharynx, and larynx, which are not rigidly supported by bone or cartilage, support is provided by muscular activity. The activity of upper airway dilator muscles is finely tuned by neural reflexes [13-17]. The laryngeal mucosa has a rich supply of receptors that are sensitive to pressure, and airflow (temperature sensitive) [13]. Activation of these receptors in the upper airway increases neural traffic in afferent nerves, primarily located in the superior laryngeal branch of the vagus nerve. Increased afferent nerve activity in turn activates upper airway dilator muscles so that muscle tension is appropriate to counteract collapsing pressures and develops at the correct time in the respiratory cycle, i.e., during inhalation. These reflexes are markedly reduced by section of the superior laryngeal branch of the vagus nerve or by topical anesthesia of the laryngeal mucosa [13,18]. Indeed, in horses, topical anesthesia of the laryngeal mucosa increases upper airway impedance and causes various degrees of pharyngeal collapse [19].
The muscles controlling the external nares of the horse demonstrate how upper airway musculature supports the upper airway on inhalation especially during exercise. In exercising horses on inhalation, muscles dilate the external nares and enlarge the opening maximally. On exhalation, these muscles relax even to the point that in many horses tissues are flaccid and a typical expiratory flutter can be heard. Similarly, other dilator muscles in the upper airway contract on inhalation and relax on exhalation. One of the most important dilator muscles in the upper airway is the cricoarytenoideus dorsalis muscle, which serves to abduct the arytenoid cartilages and vocal folds. In horses with laryngeal hemiplegia, the recurrent laryngeal nerve (branch of tenth cranial nerve) is dysfunctional, preventing normal abduction of the arytenoid cartilage [20,21]. In this condition, the larynx fails to stay dilated on inhalation and dynamic collapse of the structure occurs. Because upper airway pressure changes are insufficient to cause dynamic collapse in resting horses, clinical signs are not apparent then; however, during exercise, when the pressure changes are large, dynamic collapse of the affected arytenoid cartilage occurs, and upper airway obstruction results [6]. Surgical treatment is aimed at stabilizing this arytenoid cartilage, thus maintaining a rigid, dilated rima glottidis [5].
Dorsal displacement of the soft palate is a common condition of exercising horses [22] and like laryngeal hemiplegia, the condition is caused by neuromuscular dysfunction [23]. Support of the normal soft palate is provided by a group of muscles including the levator veli palatini, tensor veli palatini, palatinus, and palatopharyngeus muscles. The levator veli palatini muscle elevates the soft palate during activities such as swallowing, while the tensor veli palatini muscle expands the nasopharynx during inspiration by tensing the palatine aponeurosis [24]. Sectioning of the tendon of the tensor veli palatini as it inserts into the palatine aponeurosis causes the rostral half of the soft palate to become flaccid and creates mild nasopharyngeal obstruction. However, DDSP does not occur [25]. Thus, the tensor veli palatini muscle supports the rostral portion of the soft palate. The caudal portion of the soft palate is supported by the palatinus and palatopharyngeus muscles. The palatinus muscle consists of two portions that lie on either side of the midline of the soft palate, beneath the nasopharyngeal mucosa, extending caudally from the hard palate [26]. The muscles attach to the caudal aspect of the palatine aponeurosis and terminate near the caudal free margin of the soft palate. A small muscle bundle arising from the lateral aspect of each muscle continues caudo-dorsally into the palatopharyngeal arch. The palatopharyngeus muscle originates from the palatine aponeurosis and the lateral border of the palatinus muscle. It extends caudally along the lateral wall of the nasopharynx to the pharyngeal raphe, forming part of the superior constrictor muscle group [26]. Contraction of the palatinus and palatopharyngeus muscles shortens the soft palate and depresses the caudal portion toward the tongue [24,27]. Both the palatinus and palatopharyngeus muscles receive motor innervation from the pharyngeal branch of the vagus nerve. This nerve branches from the parent vagus nerve at the level of the cranial cervical ganglion and courses cranio-ventrally along the medial wall of the guttural pouch to the dorsal wall of the pharynx where it ramifies in the pharyngeal plexus. Blocking the pharyngeal branch of the vagus nerve bilaterally induces persistent DDSP in horses, implicating dysfunction of the pharyngeal branch of the vagus nerve and the palatinus and palatopharyngeus muscles in the pathogenesis of the clinical disease [23].
External Upper Airway Support
Recently there has been a great deal of interest in an equine nasal strip (FLAIR® strip) used to mechanically support the soft tissue of the nose between the nasal and incisive bone rostral to the nasoincisive notch [28]. The strip, consisting of 3 polyester springs sandwiched between two layers of material, adheres to the horse's nose. Recent studies indicate that the strip reduces upper airway impedance, (Holcombe et al., in press) during exercise, and also decreases the severity of exercise-induced pulmonary hemorrhage [28]. These findings all suggest that the nasal strip reduces upper airway tissue collapse on inhalation. This would result in decreased inspiratory impedance during exercise and less negative inspiratory pressures. Less negative inspiratory pressure may reduce the transmural pulmonary capillary pressure. Because excessive transmural pulmonary capillary pressure is responsible for exercise induced pulmonary hemorrhage, [29,30] reduction in this pressure may account for the observed decrease in exercise-induced pulmonary hemorrhage.
Effect of Head Position on the Upper Airway
In resting horses, the air entering the upper airway must turn approximately 90º to flow from the nasal passage into the trachea. That change in direction of airflow contributes to the work of breathing [31]. During exercise, the effort needed to change airflow direction is reduced by straightening of the upper airway. Straightening not only allows air a more direct route to and from the lung, it also tends to stretch and stiffen upper airway tissues, making them more rigid and more resistant to collapse. Sport horses are often forced to exercise with their heads and necks in unnatural positions. Standardbred horses are raced and trained with their head and neck in an extended position; whereas the head and neck are often flexed in sporting events such as show jumping and dressage. Head and neck extension has little effect on upper airway flow mechanics, but head and neck flexion causes upper airway obstruction. Indeed, head and neck flexion results in a significant increase in inspiratory impedance relative to the unrestrained or extended head position [31]. Thus, head and neck flexion not only makes the angle for airflow through the upper airway more acute, it also reduces the stretch of tissues necessary to prevent their collapse on inhalation. The effect of head and neck flexion on upper airway function can be a clinical problem in some performance horses. When some of these horses are exercised in the collected position, their airway becomes so obstructed that ventilation is impeded (Fig. 1).
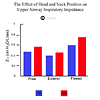
Figure 1. The effect of head and neck position on upper airway inspiratory impedance.
From the above discussion, it is clear that the upper airway has a number of mechanisms in place to prevent dynamic collapse on inhalation. The observations that upper airway resistance increases on inhalation and that a nasal strip may reduce the work of breathing suggest that these mechanisms are not totally effective. Also, upper airway lesions and physical restraints on head position may compromise upper airway dilator mechanisms.
Measurement of Upper Airway Function
The high-speed treadmill has made measurement of upper airway function during exercise practical. Evaluation of airway flow mechanics requires the measurement of airflow and driving pressure. To measure airflow, horses are fitted with a facemask and a flow measurement device. The pressures necessary to move this flow are usually measured in the mid-cervical trachea and at the external nares. The ratio of pressure and flow in a rigid tube is called resistance. However, when the walls of the tube are not rigid, like in the upper airway, and when airflow rates accelerate and decelerate, as during inhalation and exhalation, the ratio of pressure and flow is best called impedance. Measurement of air flow mechanics has been used to determine the best surgical treatments for a variety of upper airway obstructive conditions [5,6,32].
Impedance
If airflow measurements are not available, measurement of the pressure gradient along the upper airway (tran-supper airway pressure) during exercise can yield useful information [33]. As horses begin to exercise, airflow increases in response to increased trans-upper airway pressures. In upper airway obstructive diseases, the impedance provided by the airway increases and this leads either to reduced airflow or to an increase in the trans-upper airway pressures necessary to maintain flow. Increased trans-upper airway pressures by themselves have been interpreted as evidence of upper airway obstruction. Of course, this interpretation assumes that airflow rates have not decreased, an assumption that is not always correct. Nevertheless, measurement of trans-upper airway pressures, in combination with videoendoscopy, can be helpful in the determination of functional consequences of videoendoscopic findings [33].
Tidal Breathing Flow-Volume Loops
Flow-volume loop analysis is a common test of respiratory tract function in human medicine because it is non-invasive, convenient, and sensitive [34,35]. Airflow rate is continuously plotted against volume during a single, maximal inspiratory and expiratory effort. Initially used in clinical evaluation of lower airway disease only, flow-volume loop analysis has been used extensively for clinical diagnosis of upper airway obstruction in humans [36-37]. The test requires patient cooperation to perform maximum inhalation and exhalation maneuvers. In doing so, flow rates approach maximum and are associated with significant intraluminal pressure changes, allowing detection of subtle airway obstruction. In veterinary medicine, the non-cooperative nature of our patients has prevented clinical use of maximum flow-volume loop analysis.
Clinical evaluation of upper airway function using tidal breathing flow-volume loops (TBFVL) has been attempted in human neonates and infants who are incapable of performing a maximum voluntary respiratory effort on demand [38-41]. When breathing with less than maximum effort, such patients have lower airflow rates that are associated with lack of sensitivity of the TBFVL to detect airway obstruction. Furthermore, the great flow variability associated with TBFVL prevents strict numerical interpretation of loop indices, thus limiting its usefulness in evaluating airway obstruction. Tidal breathing flow-volume loop analysis performed in dogs and horses at rest lacks sensitivity and cannot reliably quantify less than severe obstruction [42,43]. The sensitivity of TBFVL in detecting airway obstruction is improved as flow rates increase. In horses, high-speed treadmill exercise results in near-maximal air flow rates. Therefore, we adapted TBFVL analysis for use in exercising horses and demonstrated that the technique is more sensitive than the measurement of flow mechanics in the detection of airway obstruction (Fig. 2) [9,44].

Figure 2. Tidal breathing flow-volume loops in a standing and exercising normal horse and in a horse with laryngeal hemiplegia. To view click on figure
Evaluation of Upper Airway Sounds During Exercise
Upper airway obstructions in exercising horses are often accompanied by abnormal respiratory noises. In racehorses, exercise intolerance associated with airway obstruction is the primary concern; whereas in sport horses respiratory noise is the most important problem as excess noise is annoying and is penalized in certain competitions. Respiratory noise in exercising horses can be recorded and analyzed with computer-based spectrogram programs [45-47]. Conditions such as laryngeal hemiplegia and dorsal displacement of the soft palate produce unique and recognizable spectrograms, and therefore upper airway sound analysis may become useful as a diagnostic tool [45].
The Lower Airway
Introduction
Diseases of the lower airway are very common. In young horses, viral respiratory diseases damage the mucociliary system and have effects on the regulation of mucus secretion and of smooth muscle that are poorly understood. Bouts of airway inflammation, probably induced by bacterial infections and the environmental contaminants in stables, are thought to be associated with poor performance in racing animals [48,49]. In older animals, severe airway obstruction is the result of repeated exposure to dusts and molds in both stables and pastures [50,51]. In the latter horses, alterations in mucus secretion and smooth muscle tension are clearly associated with airway inflammation.
There are excellent anatomical descriptions of the equine lower airway (the tracheobronchial tree) [52,53], which begins distal to the larynx and ends in the alveoli. Between the larynx and the terminal bronchioles, the airways are simply conducting tubes that deliver air to the gas exchange region i.e., the alveolar ducts and alveoli. Gas exchange does not occur in the conducting airways and they therefore form part of the respiratory dead space.
The tracheobronchial tree has defense mechanisms that protect the lung from injurious inhaled agents such as dusts, pollutant gases, antigens, and infectious agents. Defense mechanisms include cough, a mucociliary system and phagocytes to clear inhaled materials; smooth muscle to narrow the air passages and prevent entry of more agents; and a bronchial circulation that provides the inflammatory response of the air passages to the injurious agent. In diseases of the tracheobronchial tree, these defense mechanisms are employed to varying degrees, but ultimately airway diseases narrow the air passages, leading to more difficult breathing and to less efficient gas exchange. This in turn impairs the horse's performance.
Functional Anatomy of the Airways
The lower airway consists of a series of branching tubes. In the horse, the branching pattern is monopodial, i.e., at each branch point, the daughter airway is much smaller than its parent, which progresses almost directly to the periphery of the lung [53]. All the airways are lined by a mucous membrane, consisting of the epithelium and lamina propria, under which are varying amounts of smooth muscle and cartilage. In the trachea and bronchi, epithelium consists primarily of ciliated cells and goblet cells that produce mucus. In the bronchioles there are normally no goblet cells. In this region, the secretory cell is the Clara cell [52]. At all levels of the airways, the lamina propria contains a variety of sensory and motor nerves [54-56] and blood vessels from the bronchial circulation [57]. These vessels provide nutrients to the airway wall, aid in heating and humidifying air, and, in the case of airway disease, participate in the inflammatory response. There is smooth muscle in the airway wall from the trachea to the alveoli where smooth muscle fibers form sphincters around the mouth of each alveolus. Intrapulmonary airways are surrounded by and attached to the alveolar septa, so that their diameter is affected by the pull of the alveolar walls as they are stretched during inhalation.
Air Flow, Resistance, and Driving Pressure
During inhalation, the horse’s respiratory muscles work to stretch and enlarge the lung and to move air through the air passages. Flow of air is opposed by the frictional resistance of the airways, which depends largely on their diameter; a large diameter airway having a lower resistance than a small one. Individually, a mainstem bronchus has a larger diameter and a lower resistance than a bronchiole. However, there are only two main bronchi but thousands of bronchioles in parallel. This has several important consequences. First, the large bronchi and trachea constitute the largest fraction of the airway resistance while bronchioles normally provide very little resistance to flow. Second, airflow velocity is high and flow is therefore turbulent in the large airways. In contrast, velocity is low and flow is laminar in bronchioles. Third, because airway sounds reflect turbulent airflow, most of the lung sounds normally originate from the larger airways (Fig. 3).

Figure 3. Functional consequences of airway anatomy. To view click on figure
The effort being exerted by the respiratory muscles against the elasticity of the lung and the resistance of the airways is evaluated by measuring the change in pleural pressure during breathing (ΔPpl) by use of an esophageal balloon [58,59]. In the resting horse with healthy lungs, ΔPpl reaches a maximum (ΔPplmax) of up to 10 cm H2O. During exercise, ΔPplmax increases because the horse must move greater volumes of air in less time through the airways. Accelerating and moving this higher flow requires more effort [60]. When the airways are narrowed by disease, ΔPplmax increases because airway resistance is greater.
Measurement of ΔPplmax is a useful and relatively simple lung function test [59]. When airways are obstructed or when the lung is stiffened by disease, the horse must generate a greater ΔPplmax to keep the air moving in and out of the lung. Airway resistance (RL) and dynamic compliance (Cdyn) are other measures of airway function that can be made in horses [61]. Airway resistance reflects especially the function of the larger airways, whereas Cdyn or its inverse, dynamic elastance is more indicative of peripheral airway diameter. An increase in RL and a decrease in Cdyn are primarily a result of airway narrowing, which can be a result of bronchospasm, mucus obstruction, or airway wall thickening. In many obstructive diseases, these factors coexist and have important interactions with one another. For example, an increase in wall thickness as a result of proliferation of the mucus apparatus can cause an enhanced narrowing of the airway in response to bronchospasm [62].
Regulation of Airway Smooth Muscle Contraction
In the normal animal, smooth muscle is the main regulator of airway diameter throughout the tracheobronchial tree. In the animal with airway inflammation, mucus and exudates can also obstruct the airways. In the healthy animal, smooth muscle contraction is regulated primarily by the autonomic nervous system and by some interactions with the epithelium. In disease, release of chemical mediators can cause smooth muscle contraction and airway narrowing, both by direct effects on smooth muscle and by interactions with nerves regulating that muscle [63,64].
Autonomic Nerves
The primary innervation in the trachea, bronchi, and bronchioles is provided by the parasympathetic system, which reaches the lung in the vagus (Fig. 4). When acetylcholine is released from parasympathetic nerves, it binds to M3-muscarinic receptors on airway smooth muscle [65,66].

Figure 4. Autonomic airway innervation. To view click on figure
This in turn releases calcium from intracellular stores, which causes smooth muscle contraction and bronchospasm. There does not seem to be much tonic parasympathetic activity in the horse; blockade of muscarinic receptors by atropine does not cause bronchodilation in normal animals [67]. The parasympathetic system is activated in response to inhaled threats to respiratory function. Inhaled irritants such as dusts and pollutant gases, and mediators of inflammation stimulate sensory receptors in the airway mucosa and this leads to reflex activation of the parasympathetic system. This gives rise to bronchospasm and increased mucus secretion. In horses with heaves (also called recurrent airway obstruction [68]), inflammatory mediators facilitate cholinergically-mediated bronchospasm [63]. This is why muscarinic antagonists such as atropine consistently reverse the airway obstruction of heaves [67,69-71].
Airway smooth muscle is richly supplied with β2-adrenergic receptors which, when activated by circulating epinephrine or therapeutically by specific β2-agonists, such as clenbuterol or albuterol, leads to smooth muscle relaxation [72,73]. Even though sympathetic nerves occur throughout the airways, the norepinephrine that they release is probably more important in regulating the bronchial circulation [54] than in directly regulating airway smooth muscle [65].
During exercise, bronchospasm is prevented in at least two ways. Epinephrine released from the adrenal medulla activates? β2-adrenergic receptors on airway smooth muscle. In addition, sympathetic nerves release norepinephrine that activates prejunctional α2-adrenoceptors on parasympathetic nerves. This leads to inhibition of the release of acetylcholine [74-76]. These prejunctional α2-adrenoceptors are also activated by drugs such as xylazine and detomidine. For this reason, these drugs are bronchodilators in horses with heaves [77].
The inhibitory nonadrenergic-noncholinergic systems (iNANC) is the primary inhibitory nervous system supplying the smooth muscle of the trachea and large bronchi [65]. Neurotransmission in the iNANC system involves nitric oxide [78]. but its physiological importance in the horse is presently unknown. The iNANC system is dysfunctional in horses with heaves [65,79], most likely because the increased levels of reactive oxygen species present during inflammation rapidly inactivate nitric oxide.
Nerves containing neuropeptides such as substance P also occur along the airways [56,80]. These nerves are probably more important in regulation of bronchial blood flow than in control of airway smooth muscle. substance P is a vasodilator, increases vascular permeability and attracts neutrophils. Therefore, release of neuropeptides from these sensory nerves may play an important part in the inflammatory response of the airways.
All neurotransmitters and inflammatory mediators activate intracellular signaling pathways. To read more about neurotransmitters and inflammatory mediators click here.
Bronchodilators
Knowledge of the receptors regulating airway smooth muscle and the mediators activating these receptors reveals the action of bronchodilator drugs. Anticholinergic agents such as atropine and ipratropium prevent and reverse bronchospasm by blocking muscarinic receptors [67,69,71]. In the case of smooth muscle, it is blockade of the M3 receptor that prevents cholinergically mediated smooth muscle contraction. It is important to realize that anticholinergic drugs are only effective when bronchospasm is a result of acetylcholine release. However, in horses, bronchospasm seems to be primarily mediated via the cholinergic system so anticholinergic bronchodilators are very effective [67,69,71].
The most useful β-agonists for treatment of bronchospasm are specific β2-agonists such as clenbuterol [72], albuterol [73], pirbuterol [82], formoterol [83] and salmeterol [84]. Non-specific agonists such as isoproterenol also activate β1-adrenoceptors and therefore have severe cardiovascular side effects. In general, β2-agonists have a short duration of action but newer drugs such as salmeterol have a longer duration of effect [73,84].
Methylxanthines such as theophylline are the other class of commonly used bronchodilators. They inhibit phosphodiesterase, which breaks down cAMP. The resulting accumulation of cAMP in smooth muscle causes relaxation through a variety of mechanisms. Because phosphodiesterase is abundant in almost all cell types, methylxanthines have many side effects and there is little margin of safety between the plasma levels causing bronchodilation and those causing side effects [85]. Newer agents may block the specific isoenzymes of phosphodiesterase that are important in smooth muscle, be more effective bronchodilators and have fewer side effects.
Airway Hyperresponsiveness
Nonspecific airway hyperresponsiveness is an exaggerated narrowing of airways in response to many stimuli. It is a feature of many inflammatory airway diseases and contributes to airway obstruction because the airways have an increased sensitivity to excitatory neurotransmitters, inflammatory mediators, and nonspecific irritants such as dusts. Airway hyperresponsiveness is due to facilitation of neurotransmission and of smooth muscle contraction by inflammatory mediators, and to thickening of the airway wall. Horses affected by RAO are hyperresponsive to a variety of agents including methacholine [86], histamine [87-90], (Fig. 5) and citric acid [86]. When these animals are returned to pasture, airway responsiveness wanes slowly over several weeks or months but eventually becomes like that of control animals. The latter group are unaffected by stabling [90,91].
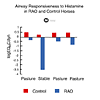
Figure 5. Airway responsiveness to histamine in RAO and control horses. To view click on figure
Airway hyperresponsiveness that persists for days can be initiated by stabling RAO-affected horses for only a few hours [92]. This observation points out the need for total environmental control so that airway inflammation is not initiated in these animals. Once initiated, a vicious cycle of inflammation, hyperresponsiveness to bronchospastic mediators, and further airway narrowing is set in motion. This cycle can only be broken by removing the horse from the initiating stimulus (usually hay dust) or by anti-inflammatory therapy [90,91].
The Mucociliary System
The mucociliary system is part of the innate defense mechanism of the lung [93]. Mucus is secreted by goblet cells and to a much lesser extent by submucosal glands in the larger airways. In the healthy horse, there are few mucus secreting cells in the bronchioles. However, in the presence of chronic disease such as RAO, mucous cell metaplasia occurs in the smaller airways [94]. Mucus consists of a protein core that is highly glycosylated. The oligosaccharide side chains account for 70 to 80 percent of the weight of mucus and confer its physical properties. Mucus is removed from the lungs by cilia, which line the airways. The rate of mucociliary clearance is greater in the larger airways and decreases toward the periphery. This arrangement prevents the accumulation of mucus as it moves from the extensive surface of the bronchioles to the much smaller surface of the trachea. When the airways are physically irritated, an increase in mucus secretion occurs very rapidly. This response is due to activation of a parasympathetic reflex. Mucus secretion also increases in the presence of inflammation. Inflammatory mediators, particularly neutrophil elastase, stimulate mucus synthesis and secretion and also cause mucus cell metaplasia. Mucus accumulates in the airways when the rate of secretion exceeds the rate of removal. One of the factors affecting the rate of removal is the viscoelasticity of mucus. In horses with acute exacerbations of RAO, mucus accumulates in the airways [95] because the production [96] and secretion of mucus is increased and also because mucus clearability is reduced by an increase in mucus viscoelasticity [97]. The increase in viscoelasticity is due to products such as DNA and actin that originate from inflammatory cells and also may be a result of alterations in the composition of the oligosaccharide side chains on the mucin molecule [95].
Pulmonary Blood Flow
The lung receives 2 circulations. The majority of the blood flow is involved in gas exchange and originates from the right ventricle via the pulmonary artery. This pulmonary circulation delivers the whole output of the right ventricle into the pulmonary capillaries. The bronchial circulation, a branch of the systemic circulation, provides nutrient blood flow to airways, large blood vessels, and the pleura [57]. The bronchial circulation can proliferate in areas of inflammation, particularly in the walls of the air passages [98].
Table 1. Vascular pressures, cardiac output and vascular resistance in the pulmonary circulation of the horse at rest and during intense exercise. Data compiled from Manohar and Goetz (1998 and 1999) [99,100]. | ||
| Rest | Strenuous Exercise |
Cardiac output (ml/min/kg bw) | 81.7 +/- 6.0 | 662.3 +/- 47.9 |
Mean pulmonary arterial pressure (mm Hg) | 31.3 +/- 1.2 | 118.1 +/- 2.5 |
Capillary pressure (mm Hg) | 27.4 +/- 1.1 | 99.3 +/- 2.6 |
Pulmonary wedge pressure (mm Hg) | 23.7 +/- 1.1 | 80.4 +/- 2.9 |
Approximate driving pressure (mm Hg) (Arterial pressure minus Pulmonary Wedge pressure) | 7 | 38 |
Pulmonary vascular resistance (mm Hg/ml/min/kg) | 0.144 +/- 0.007 | 0.060 +/- 0.010 |
Compared to the systemic circulation, the pulmonary circulation has a low resistance to blood flow. In the resting horse, mean pulmonary arterial pressure averages about 31 mm Hg (Table 1) and pulmonary wedge pressure (an approximation of left atrial pressure) is about 24 mm Hg. Hence the pressure to drive blood through the pulmonary circulation is 31 - 24 = 7 mm Hg. During intense exercise, cardiac output increases and there is a very large increase in both pulmonary arterial (118 mm Hg) and pulmonary wedge (80 mm Hg) pressures. Driving pressure for blood flow during intense exercise increases to 118 - 80 = 38 mm Hg, which is a 5-fold increase over that in the resting animal. The fact that a 5-fold increase in driving pressure accompanies an 8-fold increase in cardiac output indicates that the pulmonary circulation dilates and its resistance decreases when the horse exercises (Table 1). Dilation of the pulmonary circulation is most likely passive due to the high intravascular pressure. A role for nitric oxide has been suggested [102] but others have been unable to confirm this [99].
Because pressure increases greatly in both the pulmonary artery and left atrium during exercise, capillary pressure also increases greatly (Table 1). This increase in capillary pressure is sufficient to lead to stress failure of the capillaries [102,103] and leakage of red blood cells from the circulation into the lung. This is the cause of exercise-induced pulmonary hemorrhage (EIPH) [104] (Fig. 6). Exercise-induced pulmonary hemorrhage occurs primarily in the dorsal caudal parts of the lung [105]. This may be because these regions of the lung receive the highest blood flow [106,107] and therefore the capillaries are most distended. The negative pressure that exists in the alveoli during inhalation exaggerates EIPH. High capillary pressures and low alveolar pressures during inhalation combine to produce large transmural pressures across the capillary wall resulting in capillary wall rupture and EIPH.
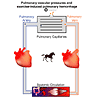
Figure 6. Pulmonary vascular pressures in standing and exercising horses and the mechanism of EIPH. To view click on figure
Very little is known about the regulation of the pulmonary circulation in horses. Nitric oxide is a short-lived vasodilator that is synthesized and released by the endothelium in response to a variety of stimuli including increased blood flow. There is no doubt that nitric oxide can dilate pulmonary arteries and veins in horses [100,108-110] but the physiological role for nitric oxide as a vasodilator during exercise is disputed [99,101]. Regional differences in endothelial nitric oxide production may be a factor in regulation of the distribution of blood flow within the lung [111] and may explain why blood flow tends to be higher in the dorsocaudal region of the lung [106,107]. Endothelin, a constrictor of pulmonary arteries may also play a role in blood flow distribution [112].
An important constrictor of the pulmonary circulation is alveolar hypoxia. This response may be useful to redirect blood flow away from poorly ventilated regions of the lung toward better-ventilated regions in order to optimize gas exchange. However, in the presence of diffuse lung disease and diffuse alveolar hypoxia, generalized constriction of pulmonary arteries can lead to pulmonary hypertension. An acute increase in pulmonary arterial pressure occurs when horses breathe a hypoxic gas mixture [113] and chronic pulmonary hypertension has been described in horses with RAO [114].
The pulmonary circulation continually filters fluid into the interstitium of the lung and, under normal conditions; this fluid is removed by pulmonary lymphatics. Edema accumulates in the lung whenever the rate of fluid filtration exceeds the rate of removal. The very high pulmonary capillary pressures that occur in the exercising horse should increase fluid filtration greatly. However, a measurable increase in extravascular lung water has not been detected in exercising horses [115]. This suggests either that the lymphatics can remove water as fast as it is filtered or that the technique could not detect the accumulations of edema fluid that occur during exercise.
Pulmonary Gas Exchange
The ultimate purpose of the lung is to exchange oxygen and carbon dioxide between the air and the blood. This requires delivery of air into the alveoli (alveolar ventilation), and the bringing together of air and blood in appropriate quantities for optimal gas exchange (matching of ventilation and blood flow). Both oxygen and carbon dioxide move between air and blood by the process of diffusion.
Alveolar Ventilation
Gas exchange cannot occur unless there is adequate delivery of oxygen and removal of carbon dioxide from lung, that is, unless there is adequate alveolar ventilation. Alveolar ventilation is regulated to keep pace with the demand for gas exchange, especially the removal of carbon dioxide. Therefore, during exercise when carbon dioxide production and oxygen consumption increase, alveolar ventilation also increases. Insufficient alveolar ventilation is known as hypoventilation and has several causes. Administration of anesthetic drugs can lead to hypoventilation by depressing the central control mechanisms that regulate ventilation. Thoracic injury, pneumothorax, or a bloated abdomen can make the efforts of the respiratory muscles inadequate to maintain ventilation. Obstruction of the upper airway by a mass or as a result of neuromuscular dysfunction (for example laryngeal hemiplegia) can also cause hypoventilation especially during exercise [116]. Severe stiffening of the lung as a result of diffuse fibrosis or lack of surfactant can be another cause hypoventilation. In horses with the diffuse airway obstruction characteristic of heaves (RAO), there can be very modest hypoventilation but severe hypoventilation does not occur even in extreme cases of the disease [71,117]. Alveolar hypoventilation is diagnosed by an increase in PaCO2 and is corrected by removing the cause or by providing positive pressure ventilation.
Matching of Ventilation Blood Flow
For gas exchange to occur, air and blood must be brought into close proximity in the alveoli. Optimal matching of ventilation with blood flow throughout the lung would be ideal but this rarely occurs. Even in the normal animal, some parts of the lung get more blood flow than ventilation while others get more ventilation than blood flow [118]. In disease, the primary cause of hypoxemia is ventilation/perfusion mismatching. When there is lung disease, the normally modest mismatching of ventilation and blood flow is exaggerated because many of peripheral airways serving the diseased lung are obstructed and blood flow may be redirected to in these inflamed regions of the lung [119]. In horses with heaves, the magnitude of ventilation perfusion mismatching is correlated with the clinical signs and severity of bronchiolar epithelial hyperplasia. In these animals, the major gas exchange defect is overventilation of some poorly perfused regions of lung [120]. When horses are under anesthesia, mismatching of ventilation and blood flow occurs because the lower part of lung is unable to expand and thereby ventilate [121,122]. There is some hypoventilation [123] and mismatching of ventilation and blood flow when horses are exercising and this contributes in part to the hypoxemia of exercise [124]. However, because the transit time of the blood through the lungs is so rapid [115], the main cause of this exercise hypoxemia is failure of diffusion equilibrium [125].
Diffusion
Diffusion is the passive movement of oxygen and carbon dioxide between the alveolus and the pulmonary capillary. Diffusion occurs as a result of the difference in partial pressure of a gas between the two structures. Because of its greater solubility, carbon dioxide diffuses more readily than oxygen. For this reason, diffusion limitation primarily affects oxygen exchange. The best example of diffusion limitation in horses is exercise-associated hypoxemia [125]. When horses exercise intensely, the blood returning to lungs from the muscles contains very little oxygen and a lot of unsaturated hemoglobin. Because of the high cardiac output that occurs during exercise, this blood, which is very deficient in oxygen, flows very rapidly through the pulmonary capillaries [115]. There is insufficient time in the capillaries for oxygen equilibrium to occur between the alveolus and blood and so the blood leaving the lung and entering the systemic arteries is hypoxemic.
About
How to reference this publication (Harvard system)?
Affiliation of the authors at the time of publication
Department of Large Animal Clinical Sciences, Veterinary Medical Center, Michigan State University, East Lansing, MI, USA

Comments (0)
Ask the author
0 comments