
Sitatunga (Tragelaphus spekei)
Read
Order: Artiodactyla
Family: Bovidae
1) General Zoological Data
"Sitatungas of Africa are semiaquatic, spending most of their life in dense beds of Papyrus" (Nowak1999). Cotton (1935) also refers to their aquatic habitat and the fact that the animals are difficult to spot in the deep papyrus stands. Males are darker and larger than lighter brown females, and only the male has (spiraled) horns. The sitatunga is still widely distributed in East Africa but is said to have nearly disappeared from the west of Africa. Many animals are held in zoological gardens. Gotch (1979) indicated that the name sitatungaderives from an old Bantu language; tragos is Greek for male goat, and elaphos is a deer. Several regional subspecies have been nominated but not further delineated genetically.
The evolution of the tribe Tragelaphini has been of interest in general but to students of cytogenetics specifically. Most of their chromosome numbers are low (around 30) and they possess fusions of sex chromosomes with one specific autosome. The nyala is an exception with a much higher chromosome number (55-56). Wallace (1976, 1978) suggested that the nyala split of earliest, to be followed by speciation of the remainder of Tragelaphini. This, he suggested, was followed and enhanced by chromosomal fusions (see section on genetics below). Vrba & Schaller (2000) indicated that the first recognized tragelaphines are to be found in deposits of 6.5 MYA, having split from the bovid ancestors that are first seen 17 MYA. Matthee & Robinson (1999) studied the phylogeny with cytochrome b analysis of this tribe and came to the conclusion that all members should be assigned the genus Tragelaphus. They grouped nyala and sitatunga closely and suggested, from molecular time clock data, that these two species separated 9.5 MYA (!). Additional relevant studies by Matthee & Davis (2001) and Hassanin & Douzery (1999) provide little further evolutionary insight.
East African Sitatunga at San Diego Zoo.

Jones (1993) gives the longevity for Eastern sitatungas as 16+ years, that of Western sitatungas as 21+ years, longer than Mentis (1972) reported. That author also indicated that these animals do not have a strict seasonal breeding period. Neonates weigh around 4 kg; Singletons are the rule, although rarely twins have been observed - twice among 1087 births (Densmore, 1980). Gestation lasts between 240-250 days and was reviewed by the same author.
2) General Gestational Data
Gestation lasts 240 and 250 days, as indicated above with usually singletons born. Neonates weigh around 4 kg and twins are uncommon.
3) Implantation
Early implantational stages have not been described and little is known of timing and manner of early placental formation.
4) General Characterization of the Placenta
I have had the opportunity to study two placentas. One was from a normal term delivery of a living offspring. It weighed 350 g and had only 28 very thin cotyledons that measured between 8 and 5 cm in diameters. The implanted cotyledon, however, was 1.5 cm in maximal thickness. The umbilical cord was 10 cm long.
The other placenta was attached to the uterus following Cesarean section of a term fetus. The pregnancy was accompanied by "hydroallantois" and remained attached to the uterus; the dam died during anesthesia and following Cesarean section surgery with pleural adhesions and marked splenic hemosiderosis. The fetus weighed 3.9 kg and was normally matured. The allantois was hugely distended with a very small amnionic cavity. The cord measured 10 cm in length, as the previously described cord, but there were approximately 40 cotyledons present in this implanted placenta. The female neonate also expired. The placenta had four rows of cotyledons.
The general structure of the cotyledons is similar to that described for eland and bongo (Hradecky et al., 1987) although the number of cotyledons enumerated for eland (142, 151) and bongo (155) by Hradecky (1983) was very much smaller (28, 40). In the chapter on bongo the placentas are also recorded as having many more cotyledons than was true for the present species.
An additional placenta was received in May, 2004 and was still partially attached to a stillborn (2.45 kg); it is depicted next. This placenta had 25 large, flat cotyledons and weighed 460 g. It was similar in all respects to that of the previously described specimens.
Sitatunga placenta from stillborn fetus.
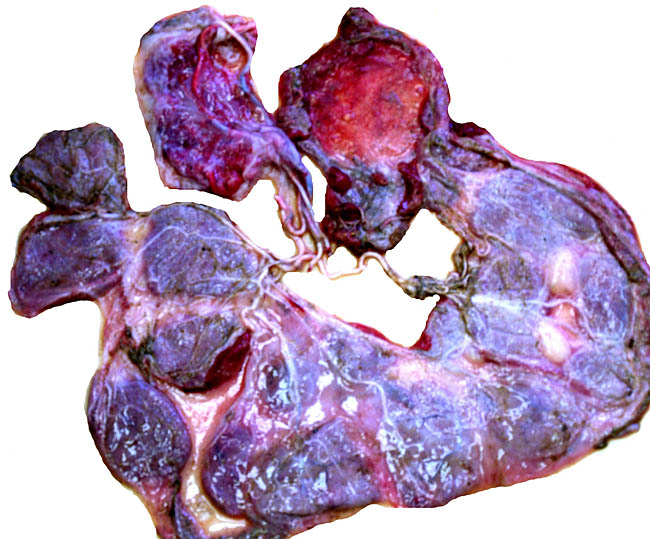
Margin of implanted cotyledon with myometrium below.

5) Details of fetal/maternal barrier
This is a typical multicotyledonary placenta with epitheliochorial contact to the uterus. There is much of the usual yellow-brown pigment accumulated beneath the chorionic plate of the cotyledons. The nature of the pigment has been discussed in various chapters on other ungulates of this web site. While it has been suggested to have a derivation from degenerating blood, it is always iron-stain-negative. I have suggested that it may be melanin deposits.
The villous surface is covered by a single layer of trophoblast among which are numerous binucleate cells. The trophoblast has intimate contact with the endometrial epithelium which is, again, single-layered. The following photographs show this well. The villi have central capillaries and scant stroma and, in my attached placenta shown, they are rather more edematous appearing beneath the chorionic plate than near the maternal aspect. They become more slender at that point.
The origin of binucleate cells that are so characteristic of ruminant placentas has been studied by numerous investigators, primarily in the sheep and cow. They are believed to produce a variety of glycoproteins, especially placental lactogen and Wooding (1982; Wooding et al., 1997) suggested that they arise by fusion of trophoblast with endometrial cells.
Surface of the placenta with few pigmented trophoblastic cells beneath chorionic plate. Above is the reflected membrane with tall trophoblast lining.
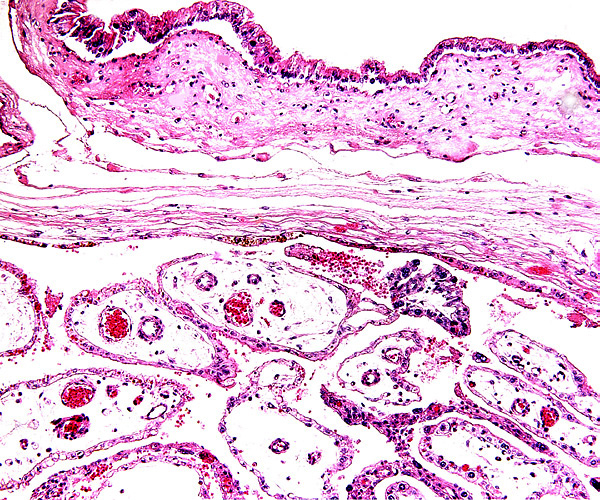
Longitudinal villus with obvious binucleate cells in endometrial glandular space.
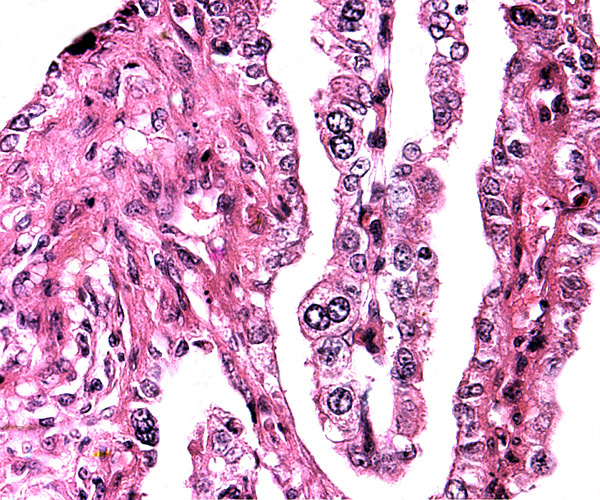
Binucleate cells top left and slender fetal villi of term placentad.
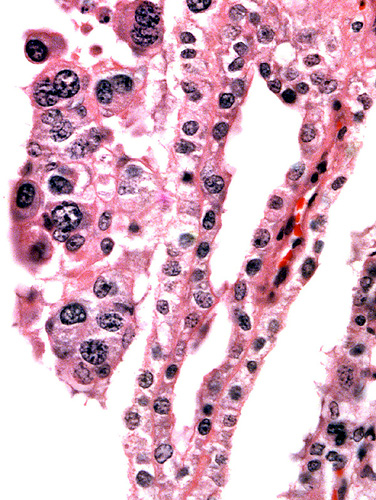
Villus in endometrial space at placental floor.
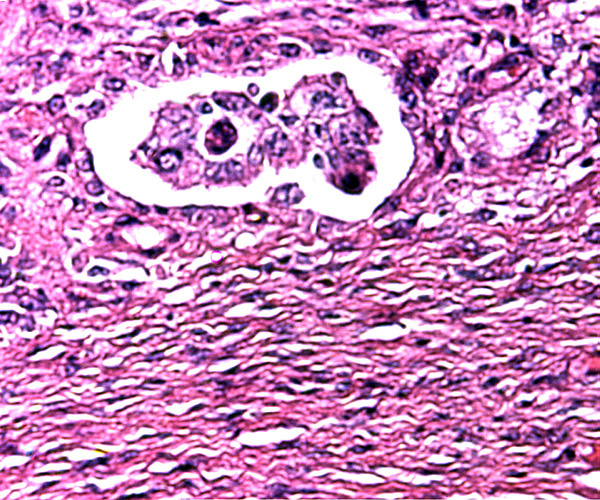
This depicts the central portion of the implanted cotyledon and is meant to show the organization of myometrium, above which are endometrial glands and dense fibrous endometrium with abundant maternal blood vessels. Follow the maternal tissue extending between villi.
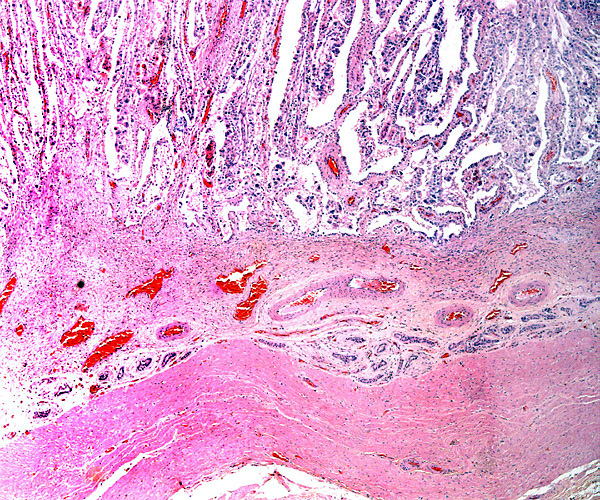
Enlargement of implantation site with interdigitation of maternal and villous tissues.
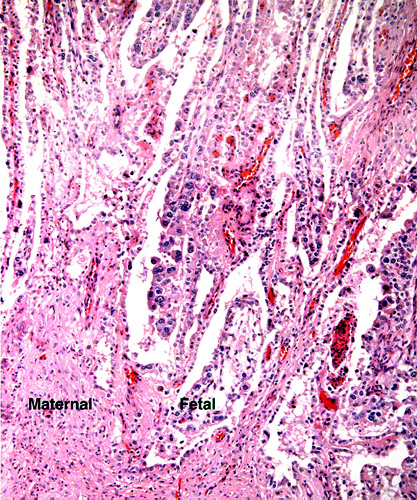
Implantation site with reining endometrial glands at bottom. The interface between villous tissue and uterus is fibrous. (M=maternal tissue; F=fetal villi).

6) Umbilical cord
Both umbilical cords were 10 cm long, unspiraled and without plaques on their surfaces. They had four large blood vessels and numerous small allantoic vessels. There was a central allantoic duct.
The cord of the third placenta, from a stillborn infant that had the placenta still attached measured 11 cm and was also unspiraled.
7) Uteroplacental circulation
I am not aware of any published studies of the vascular aspects of the bicornuate uterus in this species.
8) Extraplacental membranes
The amnion is rather smaller than the allantoic sac and is made of a single layer of flat squamous epithelium on a sheet of connective tissue that is without blood vessels. The allantoic connective tissue contains an abundant circulation and its epithelium is somewhat more prominent and, focally at least, multi-layered. I did not find remnants of the yolk sac in these two placentas.
Allanto-amnionic membranes. The vascularized allantois is below, the amnion above.
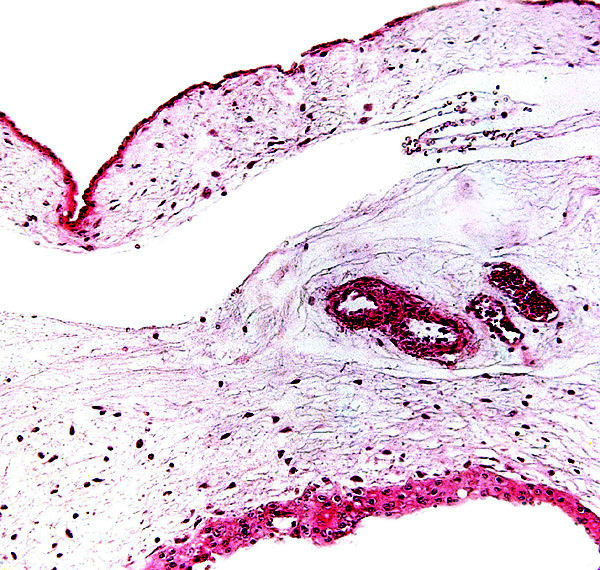
9) Trophoblast external to barrier
No trophoblastic invasion of the uterus was found in the one placenta I was able to study and that was attached to the uterus.
10) Endometrium
There are four rows of endometrial caruncles that are easily identified in the newborn uterus. The uterus is essentially bicornuate like in bovines, and not "duplex" (Hradecky, 1982). There is no classical decidua.
Cross sections through neonatal uterine horns.

Longitudinal section of neonatal uterine horn.
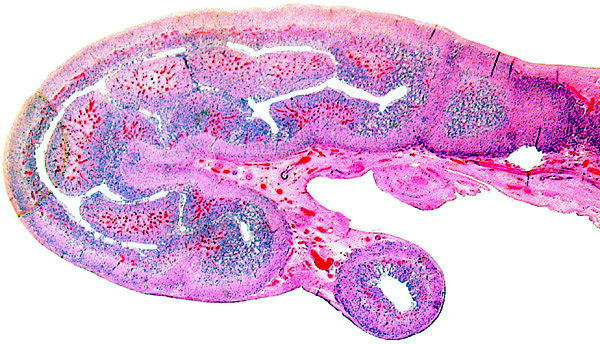
11) Various features
No other remarkable features were identified.
12) Endocrinology
The best-studied endocrine aspect of most ungulate species is the production of a variety of hormones by the binucleate trophoblast. Atkinson et al. (1993) identified a novel carbohydrate as originating from these cells and reviewed other publications that identify other glycoprotein hormones in these cells. It would appear that different populations of binucleate cells may exist with different end-products. Generally speaking, however, the cells are seen as producing primarily placental lactogen (Wooding, 1997).
No other endocrine studies of this species are known to me.
13) Genetics
Sitatungas have 30 chromosomes (Koulischer, Tijkens, & Mortelmans, 1967; Wurster & Benirschke, 1968). The sex chromosomes are unusually large - 13.08% for the X-, and 7.29% for the Y-chromosome (Wurster et al., 1968). The entire Tribe is characterized by a translocation (fusion) of the Y chromosome with an autosome (#13) (Petit et al., 1994). In addition, some members also have an autosome/X chromosome fusion (Wallace, 1976). The exception to the low chromosome number (around 30) is the nyala with 55 chromosomes (Wallace, 1978). Wallace also reviewed the possible evolution of these animals from a possible ancestor with high chromosome number and fusions following during speciation. My interpretation is depicted below as a diagram. A fertile hybrid between Bongo and Sitatunga was described by Koulischer et al. (1973). Other hybrids among tragelaphines are listed by Gray (1972) without further description of their possible fertility. The molecular studies on tragelaphines have been cited at the beginning of this chapter. They place nyala and sitatunga most closely.
Karyotype of male sitatunga.

Karyotype of female sitatunga.

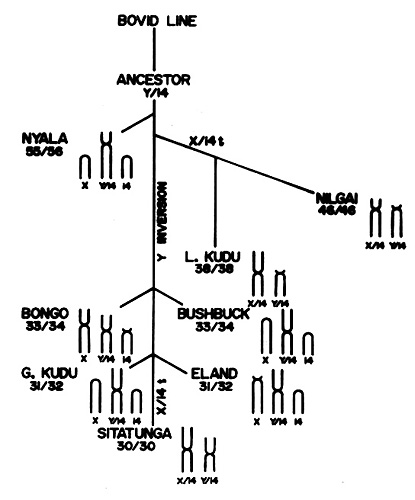
My interpretation of the phylogenetic origins of tragelaphine species.
14) Immunology
I know of no publications on immunological studies of sitatunga immunology.
15) Pathological features
In a review of neonatal mortality of 50 species of wild ungulates, Kirkwood et al. (1987) found excessively high mortality of the non-seasonal breeders, including sitatungas and attributed this mainly to management practices. Okoh et al. (1987) reported a fatal case of heartwater disease due to Cowdria ruminantium in a captive sitatunga. Walker et al. (1993) reported a new tick infection due to a novel Ixodid tick (Rhipicephalus aquatilis) in sitatungas of Tanzania, Uganda and Zambia. In a survey of artiodactyl species at Whipsnade, Flach et al. (2002) identified DNA of gamma herpesviruses in several carrier species, including the Sitatunga. A ten-year-old Sitatunga studied by Yanai et al. (2001) had calcific inclusions underneath the tongue, identified as "calcinosis circumscripta". Kaneene et al. (1985) studied the frequency of parasitic infection in ruminants kept at the Detroit zoo and were discouraged by the relatively ineffectively treated diseases in these animals.
Griner (1983) autopsied 36 sitatungas and found as main cause of death trauma and stress-related features. Four animals died from pulmonary tuberculosis due to Mycobacterium bovis.
16) Physiologic data
I am not aware of any data.
17) Other resources
A number of cell strains are kept in the "Frozen Zoo" of the Zoological Society of San Diego (CRES) and may be obtained by contacting Dr. Oliver Ryder at [email protected].
18) Other remarks - What additional Information is needed?
There is need to study the putative evolution of the tribe Tragelaphini with modern cytogenetic study, such as FISH. There is a need for the study of early implantation stages and endocrine features of gestation.
Acknowledgement
I appreciate very much the help of the pathologists at the San Diego Zoo.
Atkinson, Y.H., Gogolin-Ewens, K.J., Hounsell, E.F., Davies, M.J., Brandon, M.R. and Seamark, R.F.: Characterization of placentation-specific binucleate cell glycoprotein possessing a novel carbohydrate. J. Biol. Chem. 268:26679-26685, 1993.
Cotton, W.B.: The sitatunga (Tragelaphus spekii Sclater). Proc. Zool. Soc. London 143-144, 1935.
...About
How to reference this publication (Harvard system)?
Affiliation of the authors at the time of publication
Department of Reproductive Medicine and Pathology, School of Medecine, University of California, San Diego, CA, USA.

Comments (0)
Ask the author
0 comments