
Nilgai Antelope (Boselaphus tragocamelus)
Read
Order: Artiodactyla
Family: Bovidae
1) General Zoological Data
This large antelope, often called the blue buck, once occupied large regions of Pakistan, India and Nepal, but the range has recently contracted. Its first fossil record is from 18.5 MYA (Vrba & Schaller, 2000). The animals are frequently exhibited in zoological gardens and have done well in captivity. Since the release of 4.8 animals in Texas, a huge herd has developed there. Now, as many nilgai may live in Texas as there are left in their original habitat of Asia (Nowak, 1999). Males are darker, have short horns, a tuft on their necks, and they are larger (240-300 kg), while females are about 170 kg. Neonates weigh around 5.8 kg. First births are between 2 and 3 years of age. Longevity is 21 years and 8 months, according to Jones (1993).
Three studies of DNA have been performed to fathom the relationship of nilgai antelopes to other bovidae. Modi et al. (1996) studied repeated DNA families of 46 artiodactyl species and concluded from the findings that the subfamily Bovinae (including the Boselaphini, Bovini and Tragelaphini) was monophyletic. Matthee & Davis (2001) came to similar conclusions by studying nuclear DNA fragments. Hassanin & Douzery (1999) examined the mtDNA of the cytochrome b gene of 51 bovid species and concluded that the subfamily Bovinae includes four separate tribes, one of which is the Boselaphini.
Nilgai antelopes at San Diego Zoo.

2) General Gestational Data
Nilgai usually have twins, rarely triplets. In the United States, most are born in the fall (September/October), after mating in the preceding December. The gestation lasts 243-247 days. Estrus lasts 4 days.
3) Implantation
No studies of early implantation stages have been described.
4) General Characterization of the Placenta
The placenta of the nilgai is a polycotyledonary epithelio-chorial organ.
I have had two uteri available for study, both from the Kingsville ranch in Texas and obtained in April, and both having immature fetuses. One uterus had twins, the other a singleton. The placentas filled the entire uterine horn and there was no possible communication between placental circulations or otherwise in the twin placentas. While the cotyledons had some linear arrangement in rows, this was not so distinct as in other species and was more serpentine than I am used to seeing in ungulates. The measurements and weights were as follows:
Twins:
Length of uterine horns 21 cm; length of cervix 12 cm; weights of fetuses: female 93 g, male 84 g (14 and 13 cm length respectively). The placenta weighed 140 g after detachment. The lengths of the umbilical cords were 8 and 9 cm. There were 57 and 64 cotyledons respectively.
Singleton:
Female fetus 28 g; 10 cm length; placental weight 110 g; umbilical cord 5 cm; 65 cotyledons. The empty horn had 58 caruncles.
Pregnant uteri with immature twins.
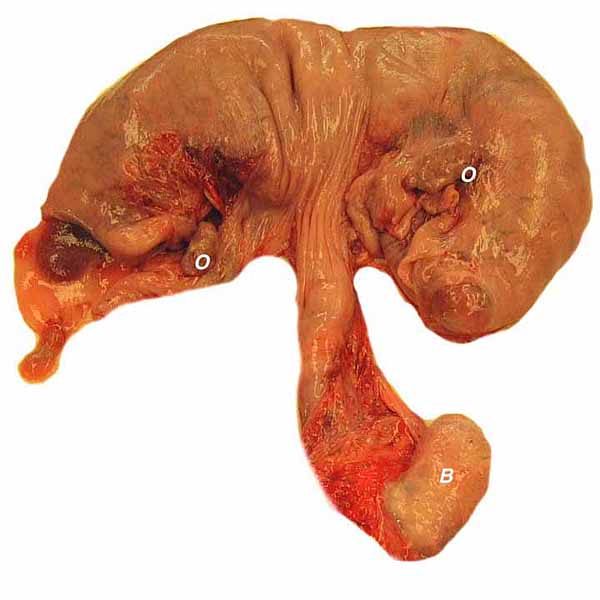
Singleton fetus with vascular uterine supply and cervices.

Immature twin fetuses in the bicornuate (duplex) uterus. Note cord insertions. Traumatic hemorrhage is seen in the left placenta.

Placenta implanted in one of the uterine horns of the twin gestation. Note the irregular arrangement of the cotyledons.
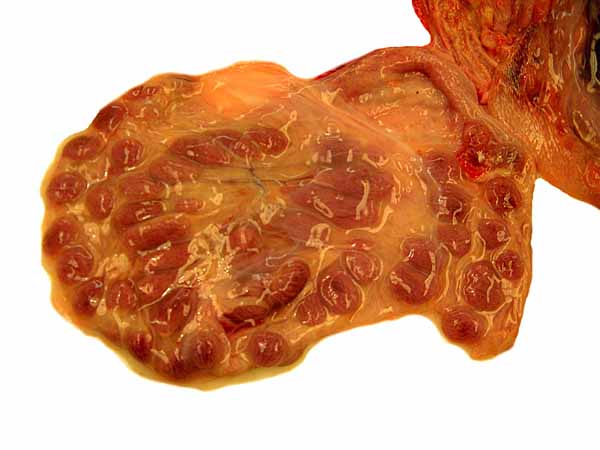
Placenta of one twin peeled from the uterine horn.

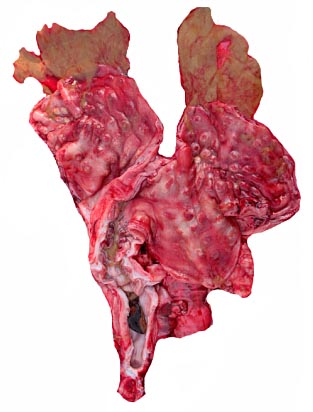
The other two uteri came from a post partum female and one from an adult but nonpregnant nulliparous dam. They are depicted below. Only in the youngest animal was there an obviously linear arrangement of the caruncles. Nevertheless, no "rows" could be enumerated and random sections through this immature uterus shows perhaps as many as six caruncles. The large size of uterine arteries supplying the endometrium is impressive. The post partum uterus contained green, mucoid secretions. The caruncles were involuted and had slightly hemorrhagic centers. Histologically, the endometrium was focally hemorrhagic and edematous. Occasional remains of detached villi were present in some glands, attended by mild lymphocytic infiltration.
Post partum uterus. This contained a mucoid, green secretion in both horns and thicker, darker mucus in the endocervix.
The uterus of the nonpregnant adult (?late juvenile) was larger on the left than on the right and had pristine caruncles that had the appearance of brownish elevations in the endometrium; they were arranged in rows.

Formalin-fixed opened uterus of immature dam to show caruncular rows.

An additional full term macerated fetus (3.5 kg, 45 cm CR length) with its placenta was available. It came from an injured dam that must have been in the process of delivery before death. The severely macerated fetus was apparently at full-term in April. This occurred in San Diego, however, where young are expected in the fall. Remarkably, one ovary had two active corpora lutea, while the other (of the nonpregnant horn's side) had a fibrotic corpus luteum. This is unlike the ovaries in the other two gestations.
5) Details of fetal/maternal barrier
This polycotyledonary placenta has all the characteristics of ruminant placentas. It is epithelio-chorial, with the villi closely interdigitating among the maternal caruncular tissues. There is a single layer of cuboidal trophoblast with numerous binucleate cells. The nature of binucleate cells and their function (production of placental lactogen) is more fully described in the chapter on sheep and urial placentas. In between the cotyledons one finds a more cylindrical trophoblast with a moderate amount of yellow pigmentation. At the edge of cotyledons, these cells are often very large ("giant cells") and their cytoplasm contains fine granules. The more immature singleton placenta has fine blood vessels in that location that are shown below. Most of these are enlarged, congested fetal capillaries. There is minimal yellow pigment in adjacent trophoblast, presumably hematoidin, although this has not been proven. The tips of the maternal septal columns that extend up to the chorionic plate show frequent degenerative changes. It is possible, but again unproven, that the pigment originates from these regions of degenerating maternal septa. This aspect is more full discussed in the chapters on urial and sheep placentas.
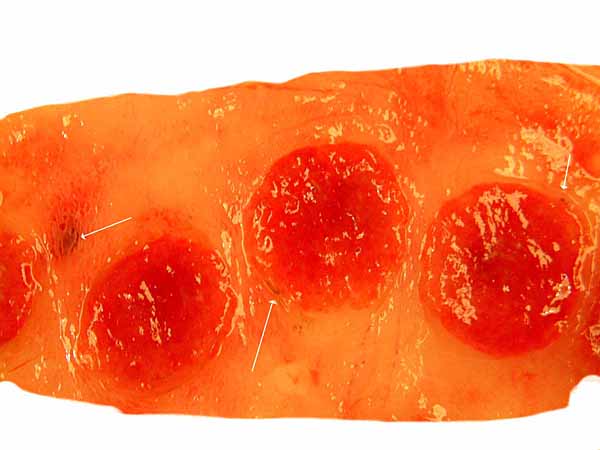
Three cotyledons of the twins' placenta (older gestation) with scant brown pigment accumulation shown at the arrows.
Edge of nilgai cotyledon with minimal pigment in trophoblast below chorionic plate. Note the degenerating material beneath.
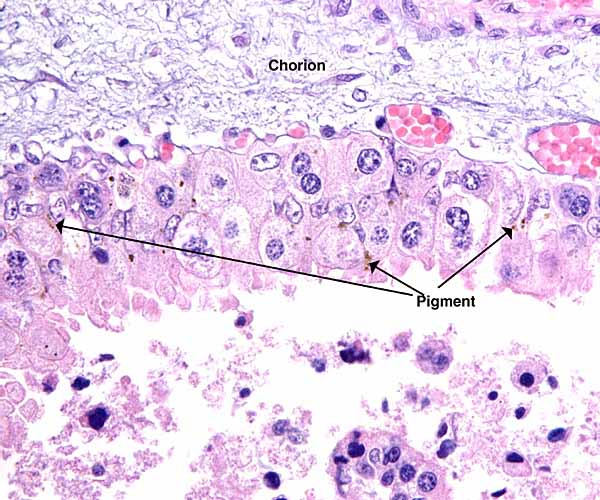
Cotyledons of the younger, singleton pregnancy with minor hemorrhages and distended vessels at the same location as the previously shown pigment deposits.

Edge of cotyledon to show the degenerating maternal tissue between tips of villi.

The same photograph, cleaned slightly to reveal the writing. B.N.=binucleate cell.

Immature nilgai placental cotyledon with its right margin fully displayed.

Two adjacent cotyledons and the umbilical cord with its central allantoic duct.

Complete cotyledon of the immature twin gestation with adjacent uterine wall.
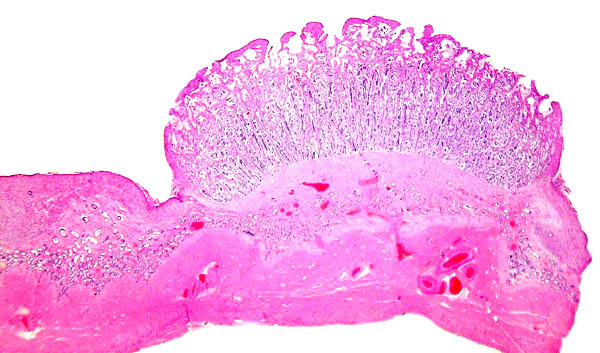
Implantation of villi within the white spaces of endometrium. They interdigitate with the endometrial caruncular epithelium but they do not invade.
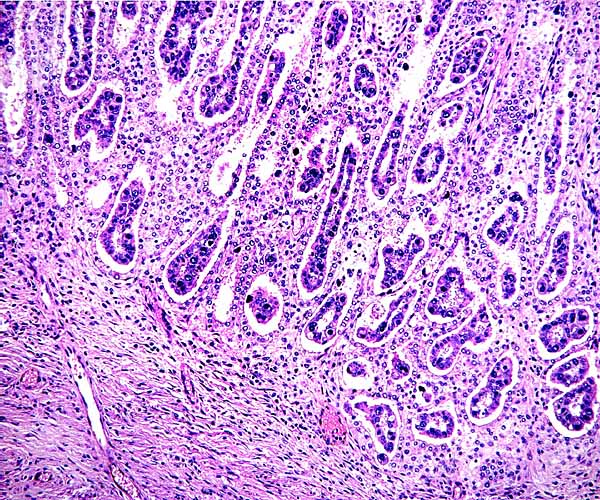
Implantation site of nilgai placenta shows a sharp delimitation of fetal and maternal cells. The trophoblast remains on the villous surfaces, and is not invasive into the fibrous endometrial stroma. Endometrial glands underlie the entire cotyledon.

Higher magnification of the villous structure of the immature nilgai placenta as it is intertwined with the maternal septa. Binucleate cell shown at arrow.

The edge of a cotyledon shows the endometrial surface and degenerative debris, some of which results in the pigmentation of the trophoblast at the edges of cotyledons.

Trophoblast below chorionic plate with huge giant cells.

Trophoblast below the chorionic plate with pigment granules and fetal blood vessels above.
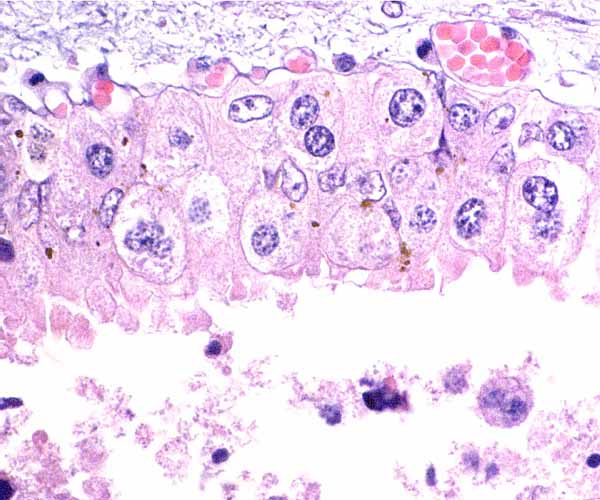
Similar view as preceding photograph.
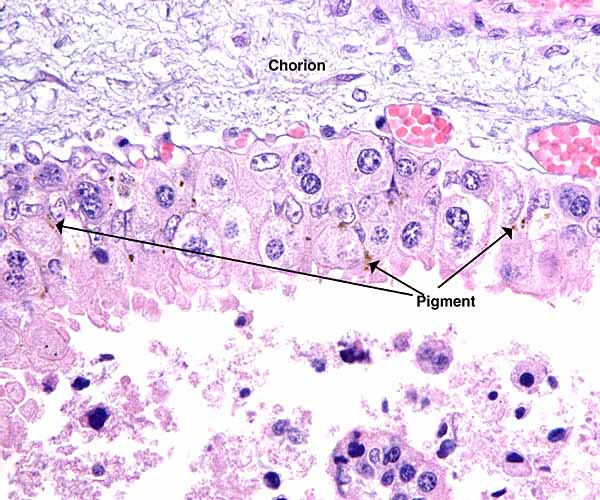
6) Umbilical cord
The umbilical cord lengths of these immature fetuses were 9, 8, 5 cm respectively. They were not coiled and were diffusely covered with fine granules. The cords contained 4 blood vessels and a central allantoic duct. The epithelium of the allantoic ducts is very thin and of urothelial type. One cord had the usual fine blood vessels, the other had none. The granules on the surface of the umbilical cord are fine squamous metaplastic foci.
The surfaces of the umbilical cords have fine granules of squamous metaplasia.
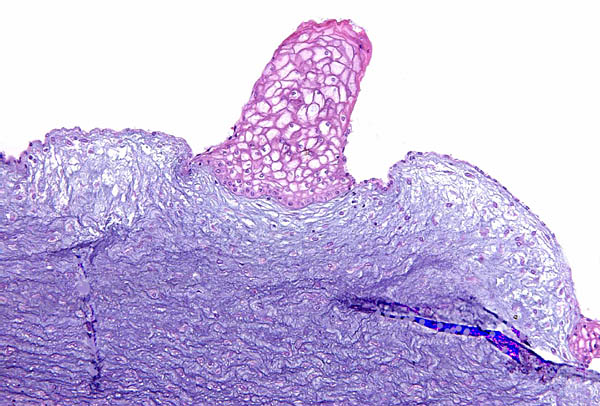
The allantoic duct of the left umbilical cord is accompanied by very small blood vessels; the one below (left) is not.
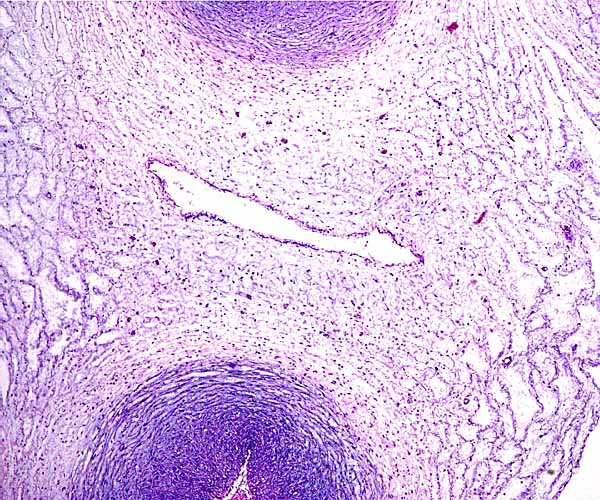
The allantoic duct of the left umbilical cord (above left) is accompanied by very small blood vessels; the one on the left is not.
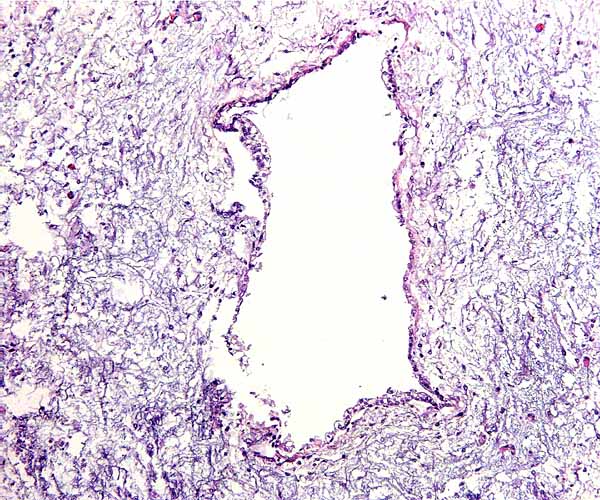
7) Uteroplacental circulation
There are no published studies, but the general architecture of the arterial arcades that supply the uterus can be seen in the singleton gestation depicted above.
8) Extraplacental membranes
This is typical of all ruminants. It is composed of an allanto-chorionic membrane whose outside is covered with cylindrical trophoblast that is often pigmented. Much degenerating decidual tissue is found between the trophoblastic outside and the endometrial surface. There is no decidua capsularis. The amnion is delicate and covered with thin squamous epithelium. The allantoic membrane is vascularized and covered with cuboidal to cylindrical epithelium. The endometrium beneath the implanting cotyledons is fibrous and represents the remains of the caruncles shown below.
9) Trophoblast external to barrier
There is no trophoblastic invasion of the endometrium.
10) Endometrium
Uterus of singleton nilgai gestation with its placenta having been peeled away. Note the small (left) non-pregnant horn and the irregular arrangement of the caruncles.

Non-pregnant horn of the singleton nilgai gestation to show the mucus-covered caruncles.

Caruncle of unstimulated horn in the singleton gestation. Note the infiltration of endometrial glands (blue) from below.

Two uterine caruncles of the nulliparous female. Note the prominent arterial supply.

11) Various features
There are neither subplacenta nor other notable features.
12) Endocrinology
Ovaries of singleton gestation. Atrophic-appearing ovary of unused horn (left) and numerous "corpora lutea" in the right ovary.

Both ovaries of the twin-bearing female had these numerous stimulated corpora. Which is the active corpus luteum could not be ascertained.
The ovary of the unstimulated side of the singleton nilgai gestation.

In the twin pregnancy, both ovaries had numerous equal-sized red "corpora lutea" without any clear differences and no obviously larger one to denote the CL of pregnancy. The ovary belonging to the immature singleton's pregnant uterine horn was similar, with many corpora, while the other, non-pregnant side was totally unstimulated. This inactivity may be the result of a diminished utero-ovarian circulation of the non-pregnant horn, as suggested by Bill Lasley (personal communication). Alternatively, it is possible that only one ovary is stimulated when singletons result, much as in cetacea and in a few other ungulates. Nevertheless, as can be seen from the preceding picture, follicular development progressing normally just not corpora lutea. Moreover, as indicated above, the macerated stillborn term gestation had two corpora lutea on the pregnant side, and a fibrotic CL on the empty side. Obviously, the hormonal milieu that leads to this ovarian histology is far from clear at this time.
The fetal ovaries and testes showed no endocrine activity.
13) Genetics
Nilgai antelopes have 46 chromosomes (Gallagher et al., 1998). Their detailed karyotypes showed many specific rearrangements when compared with the bovine karyotypes. One chromosome is apparently dicentric (Modi et al., 1996). Hybrids have not been described to my knowledge.
Petit et al. (1994) showed that tragelaphinae and boselaphinae have a Y/A translocation but that the autosome involved in this translocation differs in these two antelope lineages.
In 1941, four male and eight female nilgai antelopes were released at the King Ranch in Texas. They have proliferated enormously and there has been no evidence of inbreeding depression. Thousands are harvested annually commercially (Sheffield et al., 1983; Benirschke & Kumamoto, 1991).
Pepin et al. (1995) studied the conservation of microsatellites and their polymorphism in various species, including the nilgai antelope.
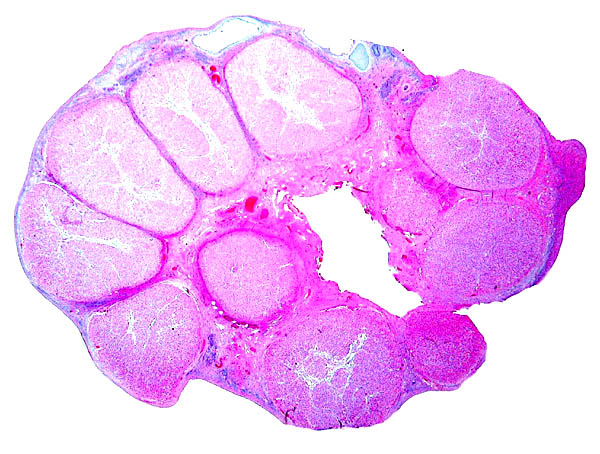
Karyotype of male nilgai antelope with 46 chromosomes.

14) Immunology
No studies have been published to my knowledge.
15) Pathological features
Rumenitis was described by Erken & Wolters (1979). Subclinical infection with the "Pestes des petits" virus was reported by Furley et al. (1987). A lymphoproliferative disease due to infection with malignant catarrhal fever virus was reported by Blake et al. (1990). The cattle tick Boophilus annulatus causes disease and mortality according to Davey (1993). Post mortem studies have shown occasional infection with Gongylonema (Chakraborty, 1994). Diverticulitis was described by Rajan et al. (1994). An outbreak of dermatomycosis due to Trichophyton mentagrophytes was described in a captive herd by Otcenasek et al. (1978). The lesions were successfully treated with antifungal agents.
Griner (1983) performed autopsies on numerous nilgai antelopes at the San Diego Zoo and found that trauma was the commonest cause of death. Two congenital anomalies were found (cardiac septal defect; joint anomaly). Thiamine deficiency was another unusual finding.
Post partum endometrium with focal hemorrhage.

Another caruncle (left) and arcade (right) of postpartum uterus with marked edema.

Postpartum endometrium with retained villi (top right) and chronic inflammatory cell infiltration.

16) Physiologic data
Priebe & Brown (1987) provided information on the nutritional requirements of growing nilgai. The structure of ribonucleases in various bovids was studied by Beintema (1980). Basic hematological values were provided by the study of Peinado et al. (1999). Hagey at el. (1997) have shown consistent changes in bile acid composition with maturation of bovids, including the nilgai.
I am fond to point out that a thick mucus plug normally occludes the endocervical canal during pregnancy of most species. It prevents ascending infection from the vaginal canal. In the absence of this protective mucus barrier, this becomes a prevalent cause of premature births in humans.
Thick mucus (blue) normally fills the endocervical canal.

17) Other resources
Cell strains of numerous animals are available from CRES at the Zoological Society of San Diego by requesting them from Dr. Oliver Ryder at [email protected].
18) Other remarks - What additional Information is needed?
Endocrine data are virtually lacking and there is no description of implantational stages. The comparison of the morphology and other aspects of physiology, genetics, etc. suggests a closer relationship to the okapi than is commonly appreciated (see the okapi chapter).
Acknowledgement
I appreciate very much the help of the veterinarians of the King Ranch in Kingsville, Texas.
Beintema, J.J.: Primary structures of pancreatic ribonucleases from Bovidae, Impala, Thomson's gazelle, nilgai and water buffalo. Biochem. Biophys Acta. 252:89-103, 1980.
Benirschke, K. and Kumamoto, A.T.: Mammalian cytogenetics and conservation of species. J. Hered. 82187-191. 1991.
...About
How to reference this publication (Harvard system)?
Affiliation of the authors at the time of publication
Department of Reproductive Medicine and Pathology, School of Medecine, University of California, San Diego, CA, USA.

Comments (0)
Ask the author
0 comments