
Chinese Goral (Naemorhedus goral)
Read
Order: Artiodactyla
Family: Bovidae (Rupicaprini)
1) General Zoological Data
Gorals are widely distributed through Southeastern Siberia, Mongolia and most of China and Korea. Nowak (1999) stated that the proper name should be Naemorhedus, not Nemorhaedus. A species perhaps closely related to Chinese gorals is the red goral (Naemorhedus cranbrooki) that finds its home in Nepal, Tibet and Burma. Soma et al. (1994) asserted that there are 9-10 subspecies of goral, perhaps three good species. Because of their existence at high altitude and scarcity, relatively little information is available on these animals. Gorals are related to serows and other rupicaprines, but are relatively rarely exhibited in Western zoos. The chromosomal evolution of these animals suggests that speciation of the Rupicaprini took place from the Alpine chamois to the American Rocky Mountain goat, with numerous species such as goral and serow in between (see also Dolan, 1963). Naemorhedus refers to forest and goat; and goral is an eastern Indian name.
Chinese goral at San Diego Zoo.

Young Chinese goral at San Diego Zoo.

2) General Gestational Data
Adult gorals weigh around 22-35 kg and may live at least for 17 years. Newborns weigh around 2 kg. There is usually a single young (Dolan, 1963). Twins are the exception, but they have been reported. Sexual maturity is attained at 2 years. The length of gestation is said to be 250-260 days (+/- 7 to 8 months). Dolan (1963) suggested it to be 180 days, but Puschmann (1984) lists 215-245 days as the gestational length. Not much literature exists on these rare animals. Soma et al. (1994) suggested that "inbreeding depression" has affected the colony held at the Japanese Serow Center.
3) Implantation
Early implantational stages have not been described.
4) General Characterization of the Placenta
I have had one pregnant uterus available for study. The female goral had died from a malignant tumor in her neck whilst pregnant in her first trimester. The entire uterus weighed 1,700 g. The fetus weighed 400 g, and it was 24 cm in crown-rump length.
The polycotyledonary placenta had 47 cotyledons and a very large sickle-shaped allantoic sac. The placenta measured 29 x 6 cm. This is an epithelio-chorial placenta, much like that of all other bovidae.
An additional placenta weighed 200 g and had a 20 x 2.5 cm umbilical cord with four blood vessels and allantoic duct.
More recently I have obtained a term placenta which weighed 375 g, had 65 coyledons and had a 12 cm umbilical cord. It is shown next.
Delivered placenta of term goral pregnancy.

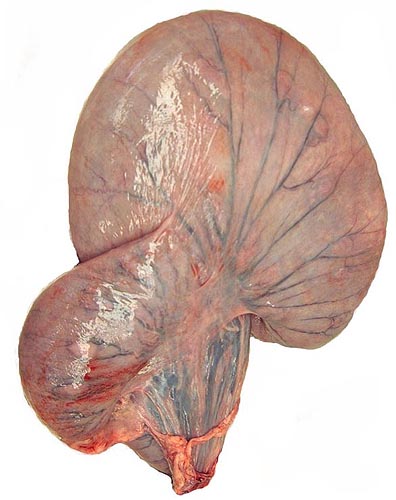
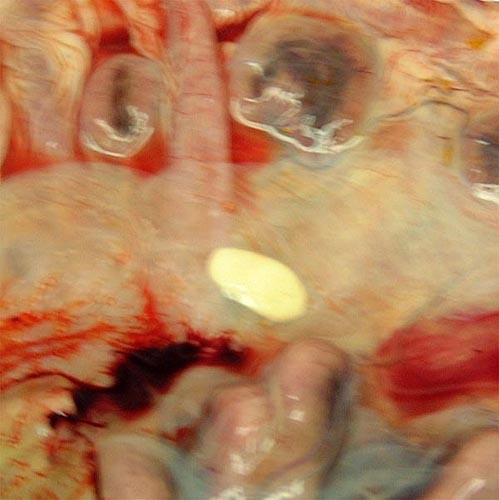
Pregnant uterus of Chinese goral.
Opened uterus with fetus. Note the brownish, large allantoic sac beneath the fetus.
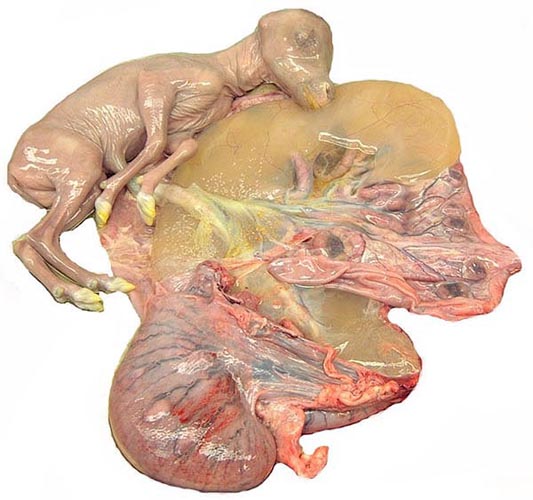
Opened uterus with the attached placenta and yellow hippomanes.
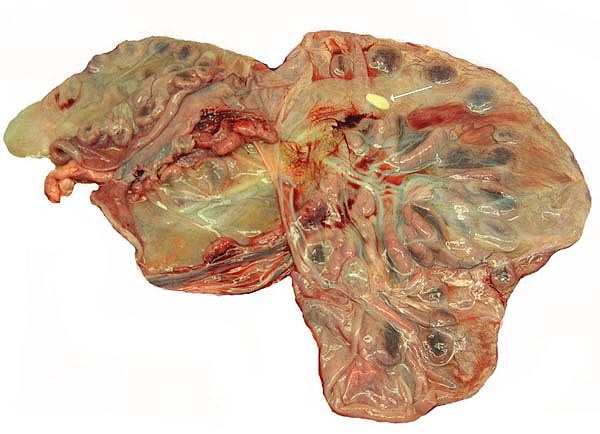
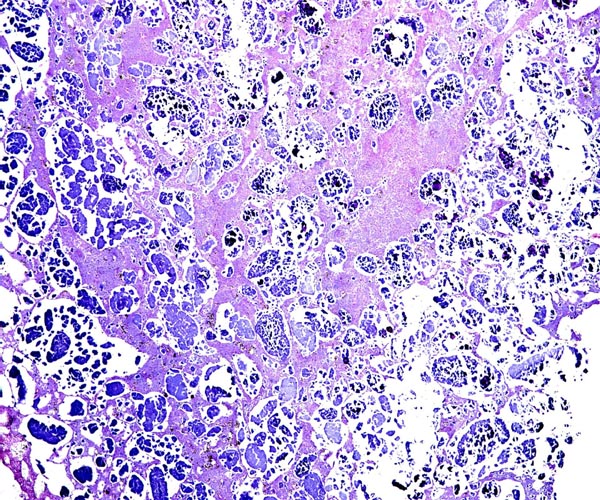
Implanted cotyledon with the myometrium below. Uterine glands are mainly located at the margins of the cotyledon.
Another cotyledon of the same goral specimen.
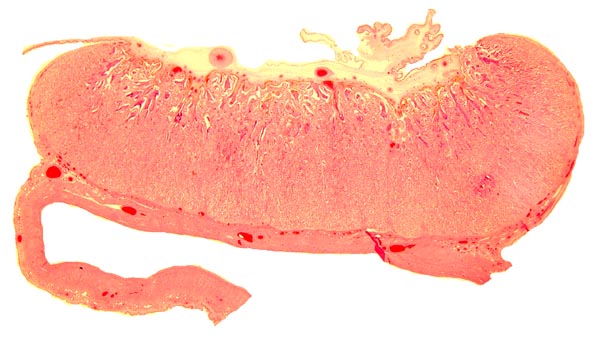
5) Details of fetal/maternal barrier
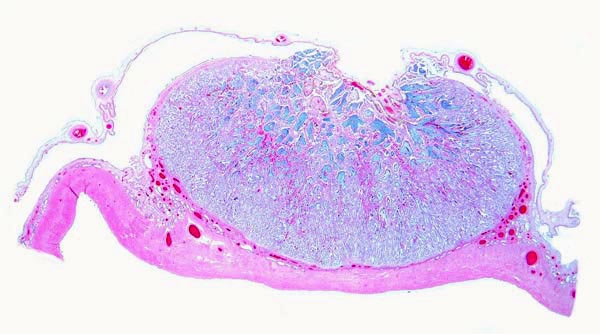
There is direct apposition of the trophoblastic epithelium to the endometrial epithelium. Invasion of the endometrium does not occur. The trophoblast is cuboidal and single-layered. Many binucleated trophoblastic cells are present and found anywhere over the villous surface. The connective tissue of the villi is scant and appeared edematous under the chorionic surface because of autolysis. Giant cells were absent. The maternal endometrial epithelium is relatively thin adjacent to the villous surfaces. Beneath the chorionic plate, the trophoblast is darkly pigmented a brown-yellow. This is presumed to be the result of maternal blood exudation.
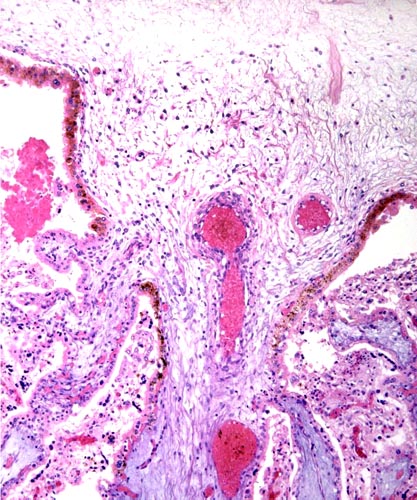
The chorion is above, and major stem villi extend downward. They are somewhat edematous (bluish) because of autolysis. Note the brownish pigmentation of the trophoblast at the undersurface of the chorionic plate.
This is the edge of a cotyledon with the villi located within the white maternal spaces, having retracted from the maternal tissues. Note the peripheral (blue) endometrial glands.

Higher magnification of villous surfaces with binucleated cells (Bi).

6) Umbilical cord
The umbilical cord was 9 cm long and had numerous fine granules on its surface. It possessed two arteries, two veins, a large allantoic duct, and numerous smaller blood vessels mainly surrounding the allantoic duct. The cord was not spiraled. The allantoic duct epithelium was thin and of urothelial type. An additional specimen had a 20 cm long cord.
Umbilical cord (arteries above) and allantoic duct in the center between the arteries. Note the superficial squamous nodules.

Surface of umbilical cord with focus of squamous metaplasia.
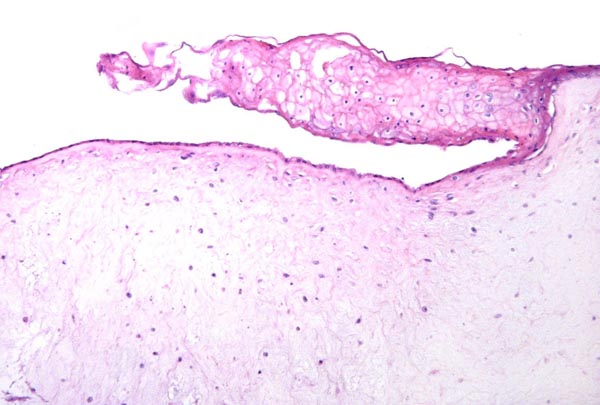
Allantoic duct of umbilical cord with adjacent small blood vessels.
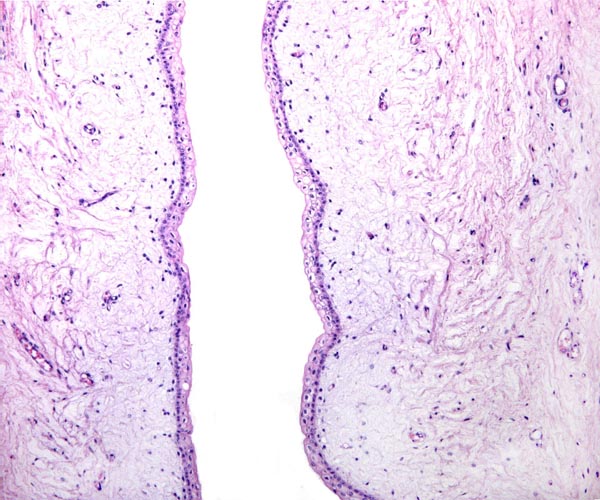
7) Uteroplacental circulation
This has not been studied.
8) Extraplacental membranes
The allantoic sac was filled with cloudy, yellow urine and had one small, deep yellow hippomanes. The epithelium is flat to columnar, especially on the intercotyledonary stretches.
Higher magnification of yellow hippomanes.
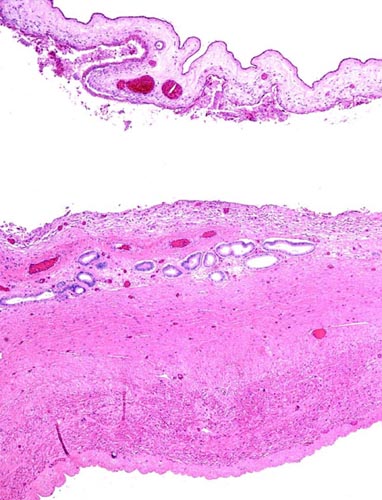
Portion of the hippomanes with calcium deposits and numerous birefringent crystals.
Surface of amnion with squamous nodule at left and allantois at right.

Myometrium and endometrium between the cotyledons. Many endometrial glands are present;the chorioallantoic membrane is at top and appears detached. This area would correspond to an "areola".
9) Trophoblast external to barrier
There is no extravillous trophoblast or invasion of the endometrium.
10) Endometrium
No true decidua is formed. The endometrial tissue becomes fibrous in appearance beneath the cotyledon and under the intercotyledonary membrane. Small numbers of lymphocytes are found within the stroma. Glands are present only at the margin of cotyledons and in between the cotyledons. None remain at the base of the cotyledon. There are markedly distended endometrial blood vessels at the cotyledonary peripheries.
11) Various features
There is no subplacenta, and no metrial glands exist.
12) Endocrinology
Melengestrol application in food has been effective in contracepting gorals temporarily (Patton et al., 2000). Cessation of the medication reversed the effect. I have not been able to find any literature on hormonal studies of orals or their closer relatives.
13) Genetics
The Chinese goral chromosome number is 2n=56 (Soma et al., 1980), all elements beings acrocentric chromosomes. In contrast, the Japanese serow has a diploid number of 2n=50, with 10 metacentrics, presumably derived by Robertsonian fusion (Benirschke et al., 1972), and the Sumatran serow with 2n=46 (Fischer & Hohn, 1972). In personal communications I learned from Dr. Shi, L of Kunming, China, that they had examined a western Chinese goral (N. g. griseus) with 2n=54 that had two metacentric chromosomes, obviously derived from Robertsonian fusion of acrocentrics. This finding helps to explain the one karyotype of 2=55 by Wurster (1972), as it may be that that specimen represented a hybrid between animals with 2n=56 and 2n=54.
Hybrids between "Grey" and Korean gorals were reported by Gray (1972).
14) Immunology
I am not aware of any studies conducted in this species.
15) Pathological features
Griner (1983) autopsied only one animal; it died with "lumpy jaws".
16) Physiologic data
There are no published data.
17) Other resources
Cell strains are available from CRES at the Zoological Society of San Diego by contacting Dr. O. Ryder at: oryder@ucsd.edu.
18) Other remarks - What additional Information is needed?
It would obviously be of interest to have some term, delivered placentas available and specimens from related species as well. Endocrine studies are completely lacking.
Acknowledgement
The animal photographs in this chapter come from the Zoological Society of San Diego. I appreciate also very much the help of the pathologists at the San Diego Zoo.
Benirschke, K., Soma, H. and Ito, T.: The chromosomes of the Japanese serow, Capricornis crispus (Temminck). Proceed. Japan Acad. 48:608-612, 1972.
Dolan, J.M.: Beitrag zur systematischen Gliederung des Tribus Rupicaprini Simpson, 1945. Z. zool. Syst. Evolutionsfschg. 1:311-407, 1963.
...About
How to reference this publication (Harvard system)?
Affiliation of the authors at the time of publication
Department of Reproductive Medicine and Pathology, School of Medecine, University of California, San Diego, CA, USA.

Comments (0)
Ask the author
0 comments